近日,九游会老哥俱乐部、省部共建生物催化与酶工程国家重点实验室吕世友/赵华燕团队在The Plant Cell上发表了题为 “An ancestral role for 3-KETOACYL-COA SYNTHASE3 as a negative regulator of plant cuticular wax synthesis” 的研究论文,揭示了β-酮脂酰CoA合成酶3(KCS3)作为负调节因子与KCS6协同精细调控植物蜡质合成的新机制。湖北大学在读博士生黄浩东和山东师范大学九游会老哥俱乐部讲师杨贤鹏为该论文共同第一作者,吕世友教授和赵华燕教授为该论文的共同通讯作者,湖北大学刘莉教授和杨平仿教授也参与了该研究,湖北大学为第一单位(图1)。

图1 文章首页
表皮蜡质是覆盖在植物表面的一层疏水性脂类化合物,主要由超长链脂肪酸(very-long-chain fatty acid, VLCFAs)及其衍生物组成。作为植物和环境之间的最外层屏障,蜡质层在植物适应外界生物和非生物胁迫方面起着重要作用。β-酮脂酰辅酶A合成酶(KCS)是脂肪酸延伸复合酶体(Fatty Acid Elongase, FAE)中的成员,决定催化底物的特异性,是FAE复合体中决定合成不同链长VLCFAs的限速酶。拟南芥KCS家族包含21个成员,根据是否具有催化活性分为两种类型。一类KCS家族成员具有FAE活性(包含α, β, γ, δ和ε分支),负责VLCFAs的催化合成,其中KCS6是参与表皮蜡质合成的关键酶。另一类KCS家族成员缺乏FAE酶活性(包含ε, η和θ分支),目前这类KCS在植物中如何行使功能仍不清楚,有待阐明。
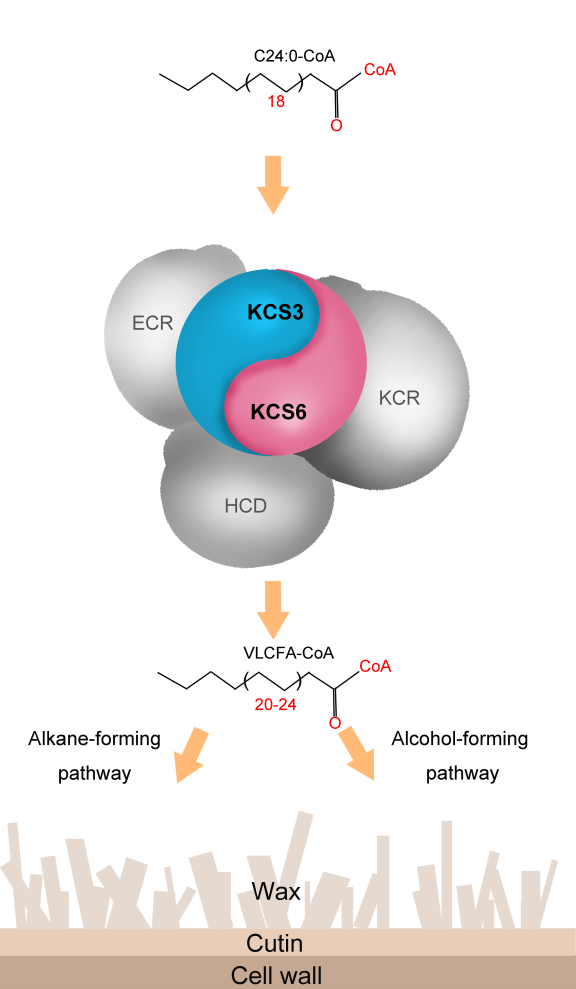
图2 KCS3-KCS6模块精细调控植物表皮蜡质合成的模式图
为了进一步拓展KCSs在蜡质合成中的作用,本研究首先构建了缺乏FAE酶活性类KCS家族成员θ分支KCS3、KCS12和KCS19在拟南芥中的过表达株系,通过蜡质组分化学分析明确了KCS3过表达能够显著抑制蜡质的合成。进一步通过遗传、异源表达分析以及荧光素酶互补成像检测和分裂泛素化酵母双杂交等实验技术手段,揭示了KCS3与KCS6互作,通过影响KCS6与其他FAE复合体成员的结合来抑制KCS6的活性,负调节蜡质合成的分子机制,并证实了该机制在古老的陆生植物苔藓中就已经出现,阐明了KCS3-KCS6调控模块在进化上的保守性。此外,该研究还发现KCS3-KCS6调控模块对维持干旱条件下的蜡质稳态起着重要作用,暗示KCS3-KCS6模块精细调控蜡质合成的分子机制在陆生植物适应性进化过程中的重要基础功能(图2)。
据悉,吕世友教授和赵华燕教授团队长期聚焦于植物脂质生物学研究,自2019年1月加盟九游会老哥俱乐部以来,以湖北大学为第一单位或通讯作者单位在Nature Communications、Plant Cell、New Phytologist和Plant Physiology等国际知名期刊或植物学经典杂志上发表10余篇高水平文章,相关研究工作得到了国家自然科学基金和湖北大学人才专项基金的支持。
论文链接: https://doi.org/10.1093/plcell/koad051



