近日,九游会老哥俱乐部、省部共建生物催化与酶工程国家重点实验室李珊珊/余希岚团队在Nature子刊(Nature Communications)上以长文形式(Article)发表了题为SESAME-catalyzed H3T11 phosphorylation inhibits Dot1-catalyzed H3K79me3 to regulate autophagy and telomere silencing的研究论文。该研究揭示了糖代谢酶特异性调控端粒结构与细胞衰老的新机制。湖北大学博士生何斐与余奇为本文的共同第一作者。湖北大学李珊珊教授与余希岚教授为本文的通讯作者,湖北大学为第一单位(图1)。

图1论文首页
端粒是染色体两端的“保护帽”,对于调控衰老非常重要。端粒具有典型的异染色质结构,有助于维持染色体的稳定性和抵抗环境压力。在酿酒酵母里,端粒结构主要由沉默信息调节因子SIR复合物来建立和维持。SIR复合物结合在端粒上可阻止转录机器结合到附近基因上,引起基因表达沉默。研究发现端粒结构受到外界营养变化、代谢水平等的调控。然而,具体的调控机制尚不清楚。
SESAME复合体是李珊珊教授在酿酒酵母里纯化、鉴定并命名的一种由糖代谢酶组成的400 kDa的蛋白酶复合体。该复合物以丙酮酸激酶Pyk1为催化亚基,通过磷酸化组蛋白H3T11(H3pT11)来抑制基因表达、调控糖代谢和增强细胞抵抗氧化压力的能力。2021年,李珊珊课题组发现SESAME复合物不仅在基因内部有结合,而且在端粒异染色质上也有明显的结合。SESAME催化的H3pT11可直接抑制自噬相关基因的表达,阻止自噬介导的Sir2降解,维持了端粒结构的完整。该课题组还发现SESAME复合物与组蛋白乙酰转移酶SAS复合物互作,特异性促进亚端粒边界区域的组蛋白H4K16乙酰化。然而,SESAME如何被特异性招募到端粒区域以及组蛋白H3T11磷酸化如何调控基因转录和染色体结构的分子机制有待进一步阐明。
为寻找H3pT11下游的效应因子,该课题组研究人员首先从H3pT11与其它组蛋白修饰在染色体上结合的相关性入手,发现H3pT11与H3K79me3在染色体分布和结合上呈显著负相关(图2)。这种负相关也出现在葡萄糖饥饿以及衰老过程中。通过体外酶反应,研究人员发现SESAME 催化的 H3pT11直接阻止 Dot1与核小体的结合从而抑制其催化H3K79me3的活性。虽然Dot1催化的H3K79me3活性被报道受H2B泛素化和H4K16乙酰化正向调控,然而负调控H3K79me3的组蛋白修饰尚不知晓。由于酵母里H3K79me3相当稳定,而且细胞里缺乏H3K79去甲基化酶,因此,该研究发现H3pT11作为首个负调控H3K79me3的组蛋白修饰,对于了解细胞精准调控H3K79me3具有重要意义。

图2. SESAME 催化的 H3pT11抑制 Dot1催化H3K79me3的活性。
有意思的是,尽管SESAME催化的H3pT11与Dot1催化的H3K79me3有拮抗关系,但是这两种组蛋白修饰协同地维持端粒异染色质结构。H3pT11分布在端粒近端,可直接招募SIR复合物到端粒区域;H3K79me3分布在端粒远端,可阻止SIR复合物从异染色质向常染色质扩散,两种组蛋白修饰联合作用可将SIR复合物稳定结合在端粒区域。形象的比喻,就像两个人运箱子,一个人在前面拉箱子,一个人在后面推箱子,同时,两个人又保持一定的距离,以免箱子被挤压坏了。进一步研究发现端粒结合蛋白 Reb1特异性招募 SESAME到端粒区域以磷酸化 H3T11并防止 H3K79me3错位结合在异染色质区域,从而建立端粒异染色质与常染色质的边界区域。当细胞在饥饿情况下,Reb1与SESAME从端粒区域解离,端粒区域H3T11磷酸化减少,H3K79me3向端粒异染色质区域延展,使得SIR复合物从端粒上脱落,使得端粒结构破坏(图3)。
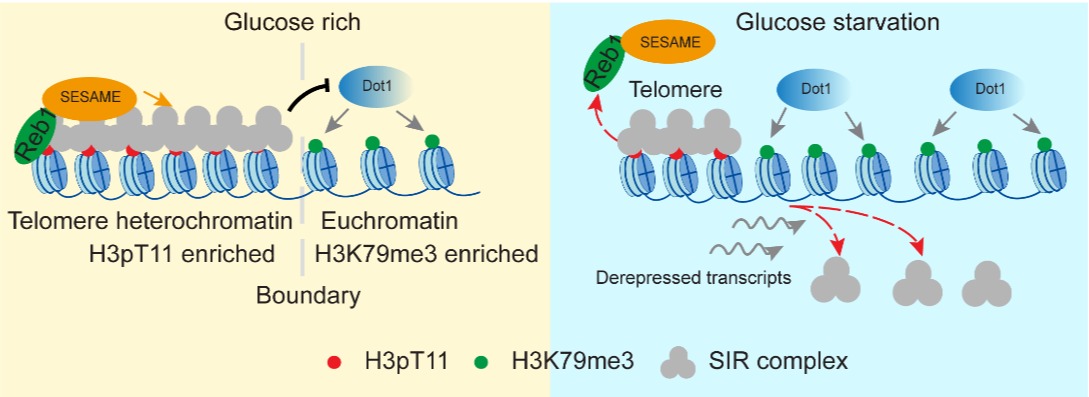
图3. 糖代谢通过Reb1-SESAME-Dot1通路调控端粒异染色质结构。
据悉,李珊珊教授和余希岚教授团队一直从事糖代谢调控衰老与肿瘤发生的表观遗传学研究。自2015年底加盟九游会老哥俱乐部以来,近5年以湖北大学为第一单位在Nature Metabolism、Nature Structural & Molecular Biology、Nature Communications (3篇)、Nucleic Acids Research (2篇)、Oncogene等国际知名权威期刊上发表20余篇高水平文章,并获得国家自然科学基金面上项目、青年项目、湖北省创新群体、湖北省杰青项目等多项国家级和省部级科研项目的支持。
原文链接:https://www.nature.com/articles/s41467-022-35182-9



